In my last blog I began a discussion on the many different DNA and RNA modifications that can modulate gene expression as part of a complex system. Over the next few blog posts I will be continuing this conversation, and today I will focus on DNA methylation.
First, what is methylation? Well, it is a structural change, namely the addition of a methyl group (CH3) to a molecule. DNA is canonically methylated on the 5’ carbon on cytosine by DNA methyltransferases. Methylated cytosines reside in CpG dinucleotides and will occur in clusters called CpG islands. Depending on how a CpG island is defined, the repeats are 200bp to 500bp in length with a majority of GC content. These CpG islands are common to promoter sites where a methyl group adds steric hindrance to the DNA strand blocking possible interacting molecules such as transcription factors or conversely interacting with methylation binding partners. Generally, DNA methylation inhibits gene expression, and ‘house-keeping’ or active genes lack methylation.
It is very well understood that gene promoter activity is controlled by transient association with a repertoire of repressors and activators. So why methylate? One possible explanation is cell specialization. How do our cells, which all have the same genetic material differentiate into kidney, bone, eye, or intestine? Can’t the transcription factors, or the chromatin structure drive these differences? They do, yet….interestingly, CpG de/methylation coincides with cell-fate decisions and development, suggesting methylation is an important mechanism driving these major cellular events. Moreover, environmental factors and lifestyle choices seem to be able to contribute to de/methylation such as oocyte maturation, embryogenesis, immune cell responses, and neural cell fate just to name a few of these life sustaining processes. Of course, one can imagine if the mechanisms that drive cell-fate decisions are disrupted, cancer could be induced. Indeed, in cancer cells the gene promoters are hypermethylated, while the gene body is hypomethylated, suggesting these regions could be therapeutic targets.
Ok, so these are some pretty remarkable scenarios regulated by methylation patterns. But the consequences of methylation can get even more wild and this I think underscores the importance of daily activities on personal health, and the well-being of our off-spring. Let me explain. Say there is a couple that wants to start a family and the soon-to-be father in the relationship eats healthily and exercises regularly. You can imagine that by doing these activities he exhibits a gene expression profile that is different than a man whom eats processed food, lays around, smokes, and drinks excessively. Now recall spermatogenesis is one of those major events I brought up earlier that involves cell differentiation and maturation. If each of these men adheres to his respective lifestyle during the time window of spermatogenesis (72 days) that produces the sperm that fertilizes the egg, methylation patterns are impaired in the unhealthy man. The healthy man does not have a gene profile or methylation profile reflective of cells coping with the consequences of smoking, lack of exercise, etc. In fact, smoking and obesity generationally transmit health consequences to offspring. Interestingly, DNA methylation is a direct link between parental health and a stunning range of disorders in offspring from cardio metabolic diseases to asthma, autism and even cancer.
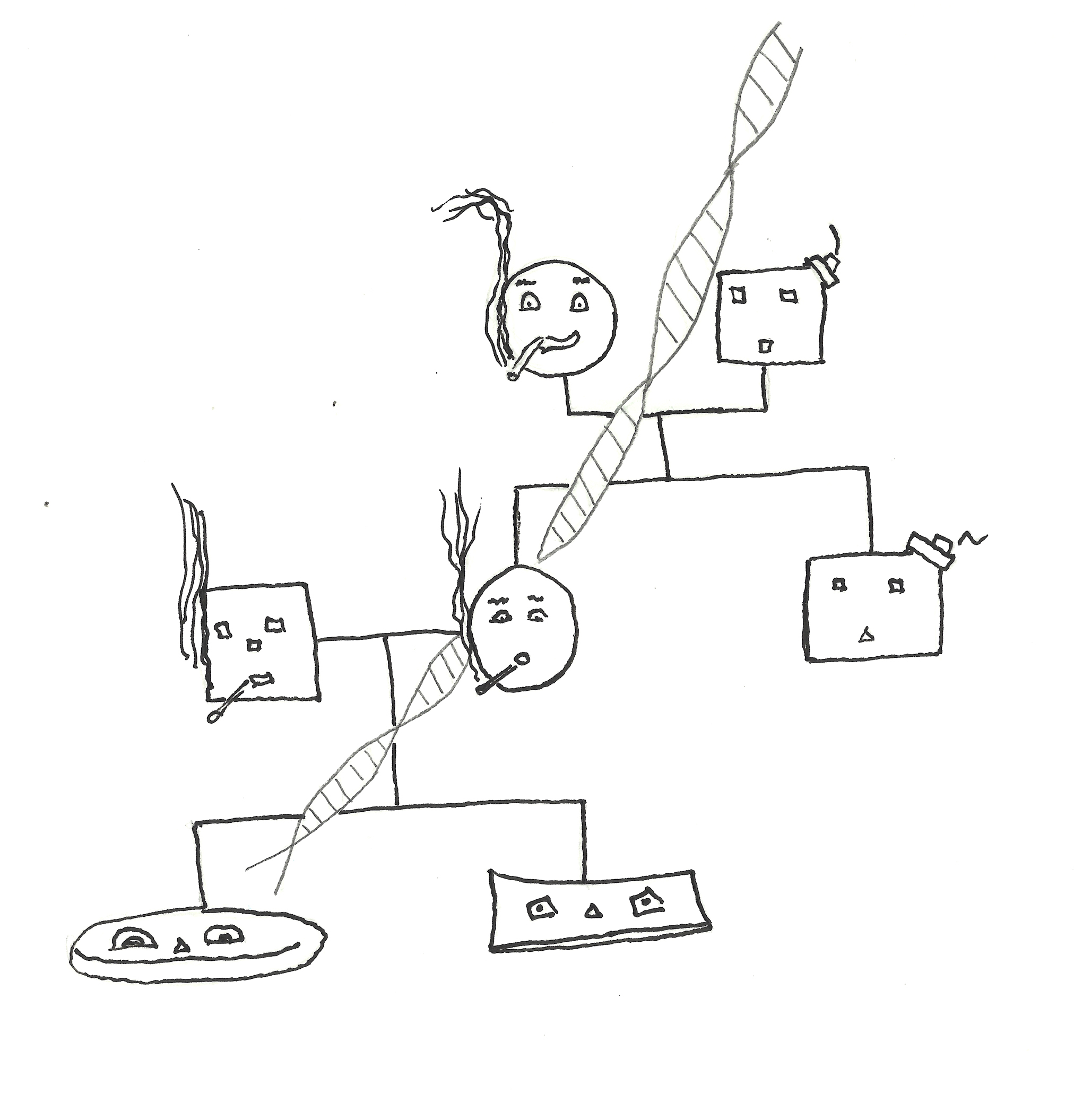
Parental habits affect future generations
So if methylation is important for gene expression, can we match patterns to definable biological outcomes? Do we need to consider for an outcome whether complete methylation of a promoter is important or rather some mosaic methylation pattern that offers the ability for fine tuning the promoter activity? If we assume that the promoter methylation mosaic is what modulates gene expression, then we open the door to some astounding numbers that suggest how complex and intricate gene expression may actually be. There are a number of different calculations on the number of CpG islands, but let’s say there are 24,000 CpG islands and each island is 200 bp in length with a hypothetical 25 CpG dinucleotides. This means for a given island there are 4.5e+47 possible methylation patterns. And for the entire human genome there would be approximately 1.08e+52 different methylation patterns. Wow. Now imagine how each of those different methylation patterns results in a different gene expression profile. Now imagine each gene profile has different transcript levels and each individual transcript has a unique function in this network. And this astounding number of possibilities culminates in a functioning circuit, the living cell. Ever since Waddington proposed the epigenetic landscape, researchers have been trying to understand the genetic and cellular parameters that define each organism on this great living planet we inhabit, Earth.
For more detail on DNA methylation and the tremendous research efforts behind our current understanding, check out the resources below as a starting point.
Resources:
Waddington, CH, The Strategy of the Genes; a Discussion of Some Aspects of Theoretical Biology, 1957, George Allen and Unwin Publishing
Robert S. Illingworth, Adrian P. Bird, CpG islands – ‘A rough guide’, FEBS Letters, Volume 583, Issue 11, 5 June 2009, Pages 1713-1720, ISSN 0014-5793, https://dx.doi.org/10.1016/j.febslet.2009.04.012. (https://www.sciencedirect.com/science/article/pii/S0014579309002919)
Uri P. Dior, Gabriella M. Lawrence, Colleen Sitlani, Daniel Enquobahrie, Orly Manor, David S. Siscovick, Yechiel Friedlander, Hagit Hochner, Parental smoking during pregnancy and offspring cardio-metabolic risk factors at ages 17 and 32, Atherosclerosis, Volume 235, Issue 2, August 2014, Pages 430-437, ISSN 0021-9150, https://dx.doi.org/10.1016/j.atherosclerosis.2014.05.937.
(https://www.sciencedirect.com/science/article/pii/S0021915014011873)
Flashner, Bess M., Russo, Mark E., Boileau, Jenine E., Leong, Derek W., Gallicano, G.Ian, Epigenetic Factors and Autism Spectrum Disorders, NeuroMolecular Medicine, Volume 15, Issue 2, 1 June 2013, Pages 339-350, https://dx.doi.org/10.1007/s12017-013-8222-5 Esteller, Manel, Epigenetics in Cancer, The New England Journal of Medicine, Volume 358, 13 March 2008, Pages 1148-1159, https://www.nejm.org/doi/full/10.1056/NEJMra072067
Soubry, A, Murphy, S K, Wang, F, Huang, Z, Vidal, A C, Fuemmeler, B F, Kurtzberg, J, Murtha, A, Jirtle, R L, Schildkraut, J M, Hoyo C, Newborns of obese parents have altered DNA methylation patterns at imprinted genes, International Journal of Obesity, 25 October 2013, https://dx.doi.org/10.1038/ijo.2013.193
Kyung E. Rhee, Suzanne Phelan, and Jeanne McCaffery, Early Determinants of Obesity: Genetic, Epigenetic, and In Utero Influences, International Journal of Pediatrics, vol. 2012, doi:10.1155/2012/463850